")
Du er her
Psykoseforståelse: et kritisk blikk på tvillingstudier
Mens det gjenstår å identifisere betydelige geneffekter for psykoser, hevdes det at tvillingstudier viser at psykoser er 70–85 % genetisk betinget. Men tvillingstudier er basert på uholdbare antakelser som gjør dem uegnet til å estimere geneffekter.

URIMELIG: Alle påvirker vi miljøet vi lever i og hvordan andre behandler oss, og vi vet at noen barn er mer utsatt for traumatiserende behandlinger enn andre barn, skriver Roar Fosse. – Men det er et skritt fra dette og til å hevde at barn selv er årsaken når de utsettes for relasjonstraumer, slik som at personer med psykose skal ha forårsaket sine egne barndomstraumer fordi de har psykosegener. Det er dette tvillingforskerne i praksis påstår.
Foto: Flikr
Skal man tro forskningen, er det liten grunn til å anta at psykoser er genetisk bestemt. Likevel holder mange fortsatt fast ved en slik antagelse, ikke minst ved å vise til tvillingstudier. Men disse studiene er lite egnet til å estimere geneffekter. Det vi derimot vet, er at alvorlig psykososialt stress er viktig. Særlig relasjonelt stress i barndommen – som seksuelt og fysisk misbruk, psykologisk misbruk, neglisjering og mobbing – er sterkt overrepresentert blant personer med schizofreni og tilgrensende psykoselidelser (Varese et al., 2012). I det siste tiåret er det også rapportert om dose-respons-sammenhenger mellom alvorlig psykososialt stress og slike psykoselidelser (heretter betegnet som psykoser) fra en rekke befolkningsstudier (Read, Bentall & Fosse, 2009). For eksempel fant en engelsk undersøkelse av 8580 personer at de som hadde erfart en type alvorlig stress i barndommen (som seksuelt overgrep eller mobbing), hadde en 1,7 ganger større risiko for psykose i voksen alder enn de som ikke hadde erfart slikt stress. For de som hadde erfart tre typer alvorlig stress, var risikoen 18 ganger høyere, og for de med fem typer var den 193 ganger høyere (Shevlin, Dorahy & Adamson, 2007).
Derimot har det vist seg vanskelig å identifisere sammenhenger mellom gener og psykoser i genetiske assosiasjonsstudier. En kritisk tolkning av denne forskningen er at ingen DNA-varianter har vist noen konsistent sammenheng med disse lidelsene. En mindre kritisk tolkning er at genvariantene som har blitt identifisert, kun forklarer en begrenset del (1–3 %) av psykoserisiko. Likevel konkluderer genforskere med at gener har stor betydning, og viser til estimater fra atferdsgenetikken som forteller at psykoser er 70 % til 85 % genetisk betinget eller arvelige (heritable) (Gershon, Alliey-Rodriguez & Liu, 2011; The International Schizophrenia Consortium, 2008). Forskjellen mellom de marginale effektene som er funnet i genassosiasjonsstudiene, og de svært høye estimatene fra atferdsgenetiske studier, beskrives i litteraturen som den manglende heritabiliteten. Denne manglende heritabiliteten har ført til omfattende diskusjoner i genfeltet. De fleste genforskere hevder at årsaken er svakheter ved de genetiske assosiasjonsstudiene, som måleproblemer, uklare definisjoner av psykoser og at DNA kan ha mer sammensatte effekter enn en tidligere har antatt. Særlig antas det at bidraget fra genetikk avhenger av samvirkningen mellom et stort antall genvarianter der hver variant har en så marginal effekt at den ikke har blitt oppdaget selv i genomvide assosiasjonsstudier med flere tusen deltakere. Med støtte i heritabilitetsestimatene brukes denne forklaringen som argument for at megastudier med titusentalls, kanskje over 100 000, deltakere er nødvendig for å påvise eksistensen av slike samvirkende gener der hvert gen har kun en marginal effekt, men hvor totaleffekten på psykoser vil være stor (Kim, Zerwas, Trace & Sullivan, 2011). En konklusjon som derimot er i tråd med funnene om manglende geneffekter, er at psykoser i liten grad er genetisk bestemt. For den svært omfattende forskningen som allerede er gjennomført burde ha funnet genene for psykoser hvis slike finnes.
Heritabilitet
Hypotesen om at psykoser er sterkt genetisk betinget, hviler med andre ord i all vesentlighet på atferdsgenetikkens heritabilitetsestimater. Heritabilitet defineres som andelen av variasjonen i et trekk i en befolkning eller et utvalg som kan forklares med genetikk. Heritabilitetsestimatene baserer seg på både tvilling-, adopsjons- og familiestudier, og på kombinasjoner av disse metodene. Den klart viktigste informasjonskilden er imidlertid den klassiske tvillingdesignen som fokuserer på eneggede og toeggede tvillingpar som har vokst opp sammen. Denne tvillingdesignen ble tatt i bruk I 1920-årene og er fremdeles svært utbredt. Den drar i dag veksler på nasjonale og internasjonale registre med til sammen flere hundre tusen tvillinger. Målet med mitt fagessay er å synliggjøre logikken i den klassiske tvillingmetoden i håp om at dette kan starte en nødvendig debatt om hvorvidt metoden gir holdbar informasjon om årsakene til psykoser.
Hensikten med den klassiske tvillingmetoden er å bestemme hvordan genetikk og miljø bidrar til et trekk (her: psykose). I tvillingmetoden blir betydningen av de to årsaksfaktorene vurdert med tanke på hvordan de bidrar til forskjellen mellom eneggede og toeggede tvillingpar i hvor like de er på trekket. Som regel måler metoden imidlertid hverken miljøforhold eller genetikk. I stedet brukes to antakelser som grunnlag for en ren statistisk estimering av årsaksforhold (se boks 1). Først, at toeggede tvillinger er halvparten så like genetisk (50 %) som eneggede (100 %). Dernest, at eneggede og toeggede tvillinger (av samme kjønn) erfarer et miljø som er likt i samme grad, dvs. at eneggede tvillingpar ikke erfarer et miljø som er likere enn det toeggede par erfarer. Denne siste antakelsen, betegnet som «Equal environment assumption» (EEA), er spesielt kritisk fordi tvillingforskerne bruker den til å legitimere at miljøet ikke betyr noe for utfallsmålet (Joseph, 2006, 2012). Dermed betrakter de genetikk som den eneste årsaken til forskjellen mellom eneggede par og toeggede par på hvor like de er på et trekk. Forutsetningen for at heritabilitetsestimatene er riktige, er dermed at EEA er gyldig. Er den ikke det, følger det at tvillingstudier overestimerer effekten av genetikk fordi miljøet da egentlig bidrar til forskjellen i trekklikhet. EEA regnes derfor som tvillingmetodens akilleshæl.
Vi skal se at forskningsevidensen tyder på at EEA ikke holder. Likevel hevder tvillingforskerne at EEA kan benyttes i estimeringen. Dette begrunner de med to tilleggsantakelser som begge innebærer at miljøet forstås som å være skapt av genetikken. Den første er at et likere miljø for eneggede par enn toeggede par skyldes førstnevntes likere genetikk. De hevder altså at den økte miljølikheten for eneggede par bør betraktes som en genetisk effekt, utløst av tvillingene selv. Den andre tilleggsantakelsen er at når foreldre eller andre familiemedlemmer utsetter tvillingene for relevante miljøforhold (som relasjonsstress for psykoser), så skyldes dette foreldrenes og familiemedlemmenes gener. Det hevdes altså at det er familiens gener som disponerer tvillingene for psykoser gjennom genarven, og som samtidig gjør at familiemedlemmene har en avvikende atferd som utsetter tvillingene for stress. Genene bidrar til at familiemedlemmene oppfører seg på belastende vis mot tvillingene, men dette ses som uvesentlig og som et tilfeldig korrelat til genene som tvillingene arver, mens genene utgjør den faktisk sykdomsdisponerende faktoren. Begge tilleggsantakelsene som brukes til å forsvare EEA, er imidlertid problematiske (se figur 1 side 1096 for en illustrasjon av logikken). Alt i alt er det derfor tvilsomt om tvillingmetoden gir holdbar informasjon om årsakene til psykoser.
Er antakelsen om like korrelerte miljøer (EEA) gyldig?
I de første tiårene med tvillingstudier (fra 1920-tallet og fremover) påpekte de fleste kritikere at eneggede par opplever et likere miljø enn toeggede par, og at dette forpurrer genetiske tolkninger. Innen vi nådde 1970-tallet, hadde tvillingforskere blitt enige med kritikerne om at eneggede behandles og sosialiseres til å bli likere, og at de oftere har samme venner og sosiale nettverk. Som eksempel konkluderte tvillingforskerne Scarr og Carter-Saltzman (1979, s. 528) med at «the evidence of greater environmental similarity for MZ “eneggede” than DZ “toeggede” twins is overwhelming». Likedan påpekte tvillingforskeren Einar Kringlen (1967) at eneggede erfarer en større psykologisk nærhet til hverandre og oftere en større identitetsforvirring og egofusjon enn toeggede.
Ifølge kritikere var og er den eneste rimelige reaksjonen på disse funnene å forkaste EEA (Joseph, 2006). Ettersom eneggede par opplever et likere miljø enn toeggede par, er det rimelig at dette bidrar til at eneggede er likere på en rekke trekk enn toeggede. Et eksempel er likhet i venner og sosialt nettverk. En rekke studier har vist at den større likheten for eneggede enn toeggede par i venner og sosialt nettverk bidrar til større likhet hos eneggede tvillinger i lidelser som depresjon, angst og PTSD (e.g. Horwitz, Videon, Schmitz & Davis, 2003). Basert på slike funn for psykiske lidelser advarte Horwitz et al. (2003, s. 125) om at «Some of the greater concordances between monozygotic, compared to dizygotic, twins that behavioral geneticists attribute to genomic factors might stem from the more common social environments that monozygotic twins share».
Tvillingforskerne forkastet imidlertid ikke EEA selv når det ble vist at eneggede tvillingpar erfarer miljøer som er likere enn det toeggede par erfarer. I stedet spesifiserte de EEA til kun å gjelde for miljøpåvirkninger som er relevante for det trekket man studerer, dvs. som betyr noe for utviklingen av det trekket man undersøker. Kenneth Kendler og medarbeidere (1991, s. 21) formulerte dette slik:
The traditional twin method, as well as more recent biometrical models for twin analysis, is predicated on the equal-environment assumption (EEA) - that monozygotic (MZ) and dizygotic (DZ) twins are equally correlated for their exposure to environmental influences that are of etiologic relevance to the trait under study.
Likedan hevdet Bouchard og McGue (2003, s. 9):
…the issue is not whether MZ twins experience more environmental similarity than DZ twins, but rather whether they are more likely to share trait-relevant features of their environments.
Tvillingforskerne sier altså at det ikke gjør noe om eneggede er likere enn toeggede på miljøforhold som ikke påvirker det trekket en studerer. EEA gjelder i stedet kun for miljøforhold som påvirker det aktuelle trekket. For psykoser betyr dette at psykososialt stress, som er empirisk knyttet til lidelsen, ikke er likere for eneggede enn for toeggede tvillinger. Heller ikke andre miljøforhold som kan tenkes å være knyttet til psykoser, bør være likere for eneggede, som prenatale belastninger og cannabismisbruk. En åpenbar svakhet er at tvillingforskerne ikke har spesifisert hvordan man skal gå frem for å bestemme hvilke miljøforhold som er relevante eller irrelevante for psykoser og andre trekk.
Når EEA ble spesifisert til å gjelde kun for trekkrelevante miljøerfaringer, kunne en forventet at tvillingforskerne målte og tok i betraktning alvorlig psykososialt stress i sine videre estimeringer for psykoser. Ingen tvillingstudier har imidlertid testet EEA for psykoser ved å måle relasjonsstress eller andre former for psykososialt stress. De har altså nøyd seg med å revidere og spesifisere EEA til å gjelde kun for relevante miljøforhold, men har unnlatt å undersøke om denne versjonen av antakelsen er riktig. For psykoser fremstår dermed den reviderte versjonen av EEA kun som en teoretisk formulering og som en hypotese som ikke er empirisk testet. Joseph (2006) kritiserte derfor tvillingforskerne for at de ikke er interessert i å teste sin egen grunnantakelse. I stedet virker det som de mener at denne oppgaven faller på kritikerne.
Selv om tvillingforskere ikke direkte har testet EEA for psykoser ved å måle psykososialt stress, har de likevel målt psykososialt stress i flere studier. Den vitenskapelige rammen for disse studiene har gjerne vært at tvillingforskerne har ønsket å undersøke hvorvidt stressbelastninger er et resultat av tvillingenes eller familiemedlemmenes gener, noe jeg kommer tilbake til nedenfor. I disse tvillingstudiene er det dokumentert at eneggede har likere erfaringer i oppveksten enn toeggede, med likere erfaringer når det gjelder mobbing, krangling med foreldrene, tap av venner, seksuelle overgrep, ran, å bli holdt fanget og å bli slått. For eksempel fant Ball et al. (2008) at mobbing i barndommen var sterkere korrelert for eneggede (r = 0,73) enn for toeggede tvillinger (r = 0,38). Likedan fant Dinwiddie et al., (2000) at seksuelle overgrep før 18-årsalderen var likere for eneggede (r = 0,68) enn for toeggede (r = 0,52), mens Stein, Jang, Taylor, Vernon og Livesley (2002) rapporterte at traumer som seksuelle overgrep, å bli ranet, banket opp og holdt fanget var likere for eneggede (r = 0,43) enn for toeggede tvillinger (r = 0,31). Slike funn tyder altså på at heller ikke den reviderte, trekkrelevante versjonen av EEA er gyldig for psykoser (eller andre psykiske lidelser), fordi eneggede tvillingpar utsettes for likere stressbelastninger enn toeggede tvillingpar.
Er psykososialt stress kun en genetisk sideeffekt?
Tvillingforskerne beholder imidlertid EEA for psykoser og andre psykiske lidelser selv om eneggede er funnet å ha likere miljøbelastninger enn det toeggede har. Dette begrunner de med to tilleggsantakelser. Begge disse tilleggene bruker de som argument for at korrelasjoner mellom genetikk og miljø gjør miljøet irrelevant, slik at miljøet heller bør betraktes som en effekt av genetikken.
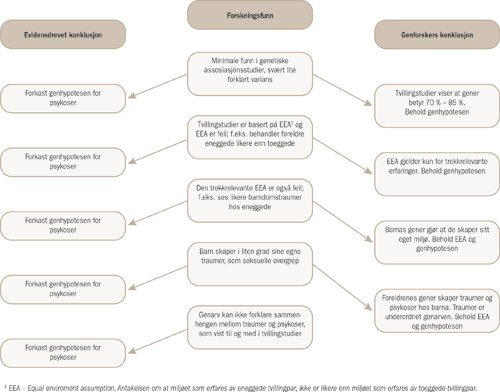
Figur 1. Spissformulering av genforskernes logikk for å beholde genhypotesen for psykoser
1 EEA – Equal enviroment assumption. Antakelsen om at miljøet som erfares av eneggede tvillingpar, ikke er likere enn miljøet som erfares av toeggede tvillingpar.
Tilleggsantakelse 1: miljøet kan ses bort fra fordi miljøet skyldes barnas genetikk
Tvillingforskere hevder at eneggede tvillinger skaper et likere miljø for seg selv enn toeggede fordi de har en likere genetikk. Dermed forstås forskjellen i miljølikhet som en genetisk effekt, og EEA kan beholdes. Som formulert av Cesarini, Johannesson, Wallace og Lichtenstein (2009, s. 621):
It is important to emphasize that even if MZ twins are treated more similarly, this does not in and of itself constitute a violation of the [EEA] assumption; greater similarity in environment may be caused by the greater similarity in genotypes.
Bouchard, Lykken, McGue, Segal og Tellegen (1990, s. 227–228) formulerte det samme poenget slik:
Twins tend to elicit, select, seek out, or create very similar effective environments and, to that extent, the impact of these experiences is counted as genetic influence.
Tvillingforskerne sikter her først og fremst til at tvillinger, som andre barn, aktivt skaper sitt eget miljø, og at måten tvillingene behandles på, dermed kan regnes som en genetisk effekt. Fordi eneggede er likere genetisk, vil de velge, søke ut, skape og utløse likere miljøer og reaksjoner fra andre mennesker enn tilfellet er med toeggede.
Det er en rimelig påstand at vi alle påvirker miljøet vi lever i og hvordan andre behandler oss. Det er også kjent at noen barn er mer utsatt for traumatiserende behandlinger enn andre barn, slik som ved mobbing. Men det er et skritt fra dette og til å hevde at barn selv er årsaken når de utsettes for relasjonstraumer, slik som at personer med psykose skal ha forårsaket sine egne barndomstraumer fordi de har psykosegener. Det er dette tvillingforskerne i praksis påstår.
For en del av oss fremstår det som absurd at ansvaret for eksempel for seksuelle overgrep skal legges til barnet selv. En slik påstand motsies også direkte av forskningen. Traumatisering av barn er empirisk knyttet til karakteristika ved utøveren, som belastninger i utøverens livshistorie, psykologiske vansker, rusmisbruk, ekteskapsproblemer, røffe oppdragelsesstrategier og avvisende tilknytningsmønstre. Likedan er mobbing utført av jevnaldrende knyttet til trekk ved mobberen og dennes livssituasjon.
I tråd med denne kritikken har tvillingforskere selv rapportert at økte miljøbelastninger i barndommen hos eneggede sammenliknet med toeggede tvillingpar ikke kan avskrives med henvisning til genetisk påvirning av miljøet. Jaffe et al. (2004) rapporterte for eksempel at genetikk ikke kunne svare for noe av variansen i fysisk mishandling i en gruppe eneggede og toeggede tvillinger og kun for 25 % av variansen i fysisk straff. Det betyr at variansen i all hovedsak må tilskrives miljøpåvirkninger. Et annet eksempel er Alemany et al. (2012), som brukte en metode der de målte forskjeller mellom eneggede tvillinger i belastninger i barndommen og i psykosesymptomer som voksen. Siden eneggede tvillinger er genetisk identiske, kan forskjeller mellom dem på et trekk ikke skyldes genetikk. Hvis så forskjellen mellom eneggede tvillinger på et trekk korrelerer med forskjeller i erfaringene deres, så viser dette at erfaringene er en årsak til trekkforskjellen. Alemany et al. (2012) fant nettopp dette for psykosesymptomer. De rapporterte at forskjellen mellom eneggede tvillinger i belastninger som overgrep og neglisjering i barndommen korrelerte med forskjellen mellom tvillingene i både positive og negative psykosesymptomer. Altså at hvis den eneggede tvillingen opplevde overgrep, men ikke den andre, økte sannsynligheten for psykosesymptomer hos førstnevnte.
Tilleggsantakelse 2: miljøet kan ses bort fra fordi det er et uvesentlig korrelat til foreldrenes genetikk
I sin tilleggsantakelse nummer to hevder tvillingforskerne at likere erfaringer hos eneggede tvillinger enn hos toeggede tvillinger kan ses bort fra i utregninger av heritabilitet, fordi deres likere erfaringer er et produkt av genarven og dermed et uvesentlig korrelat til denne. Price og Jaffee (2008, s. 305–306) beskriver dette slik:
…the family environment depends on heritable parental characteristics, so that parents pass on to their children an environment that correlates with the parental genotype… When this genotype also influences children’s behavioral or cognitive outcomes, the result is a spurious association between environment and outcome.
For eksempel, når intelligente foreldre får intelligente barn, så skyldes dette genarven, mens det har mindre å si at foreldrene (på grunn av sine «intelligensgener») samtidig gir barna et stimulerende oppvekstmiljø. Tilsvarende skyldes utviklingen av psykoser arvede gener, mens et negativt familiemiljø blir et uvesentlig korrelat som også skyldes foreldrenes gener. I en kontekst med tvillinger kan en dermed argumentere for at likere erfaringer for eneggede enn for toeggede skyldes at foreldrene tenderer til å behandle eneggede likere, men at dette er et uvesentlig korrelat til de eneggedes likere (arvede) gener. Ved å bruke dette argumentet kan EEA igjen beholdes, og det psykososiale kan fremdeles ses bort fra i estimeringen av heritabilitet. Jaffee og Price (2008, s. 496) beskriver dette slik, med antisosial atferd som eksempel:
…because parents who have histories of antisocial behavior… are at increased risk of abusing their children, maltreatment may be a marker for genetic risk that parents transmit to children rather than a causal risk factor for children’s conduct problems.
Denne antakelsen tilsier at selv de sterke dose-respons-sammenhengene som er rapportert mellom barndomstraumer og psykoser, egentlig er genetiske effekter. En må her anta at det finnes ulike doser av psykosegener som nøye samsvarer med graden av traumer som bærere av disse genene utsetter barn for. Dette vil være forbløffende, og en rekke funn tyder på at antakelsen ikke er riktig.
Først, eksperimentell dyreforskning viser at skjevutvikling i atferd skyldes miljøbelastninger heller enn genetikk. I denne forskningen har en kunnet kontrollere betingelsene ved å randomisere avkommet til biologiske foreldre og adoptivforeldre med ulik omsorgskvalitet. For rotter er det for eksempel demonstrert at det snarere er omsorgssvikt enn opphavets genetikk som forårsaker skjevutvikling i atferd og underliggende hjernefunksjon (Champagne & Curley, 2009).
Dernest kan antakelsen ikke brukes til å forklare effekten av andre typer miljøbelastninger enn relasjonsstress påført av familiemedlemmer. Eksempler er mobbing og utfrysning på skolen, overgrep på barnehjem, å være vitne til gjentatt vold i nærmiljøet og krigstraumer (Read et al., 2009). Slike erfaringer kan ikke være skapt av foreldrenes gener, og de kan derfor ikke ses bort fra på dette grunnlaget.
For det tredje, tvillingforskningen har selv vist at antakelsen er uholdbar. Ett eksempel er en studie av Arseneault et al. (2011) som fant at traumer i barndommen økte risikoen for psykosesymptomer blant eneggede og toeggede tvillinger med 322 %. Når forskerne «kontrollerte» for den større genetiske likheten mellom eneggede tvillinger, observerte de at traumer fremdeles var knyttet til en 281 % økning i psykosesymptomer. Altså kunne en (teoretisk mulig) likere arv av potensielle «sykdomsdisponerende» gener ikke forklare sammenhengen med barndomstraumer.
Antakelsen om at miljøet er et uvesentlig korrelat til sykdomsdisponerende gener, kan heller ikke forklare forskjeller eller diskordans mellom eneggede tvillinger. For psykoser vil diskordans si at kun den ene, men ikke den andre eneggede tvillingen utvikler lidelsen. Diskordans for psykose hos eneggede tvillingpar inntreffer minst like ofte som konkordans eller sammenfall (Ross, 2013). En slik diskordans kan ikke forklares med genarv, men tyder i stedet på forskjeller i miljøerfaringer mellom tvillingene og dermed på at miljøet er sentralt (Ross, 2013).
Den andre tilleggsantakelsen som tvillingforskerne har brukt når EEA har vist seg å være ugyldig, er med andre ord i strid med empiriske funn fra flere forskningsområder. Også denne tilleggsantakelsen er dermed ugyldig.
Ugyldige estimater
Antakelsen om at eneggede og toeggede tvillingpar erfarer like korrelerte miljøer (EEA), utgjør tvillingforskernes grunnpremiss for estimering av arvelighet for psykoser og andre trekk. Denne antakelsen beholdes selv når det er vist at miljøet er likere for eneggede enn for toeggede tvillinger, inklusive psykososialt stress – den mest relevante miljøfaktoren for psykoser. I stedet for å konkludere med at EEA ikke kan brukes som grunnlag for estimeringer av arvelighet, har imidlertid tvillingforskerne fremmet to tilleggsantakelser for å forsvare den fortsatte bruken av antakelsen. De hevder da at (1) eneggedes likere genetikk gjør at de skaper likere belastninger for seg selv enn toeggede, og (2) når familiemedlemmer utsetter tvillinger for traumer, så er dette underordnet effekten av genarven. Som vist her er disse antakelsene uholdbare.
Tvillingforskernes konklusjoner om at psykoser i hovedsak beror på gener, hviler altså på de samme antakelsene de alt har gjort seg i utgangspunktet (Joseph, 2012). Når EEA i tillegg viser seg ikke å holde, så setter tvillingforskerne frem andre antakelser for å forsvare den. Logikken som da brukes, viser seg å være sirkulær; antakelser om gener brukes som grunnlag for konklusjoner om gener og konklusjonene som grunnlag for antakelsene (Joseph, 2012). I slik sirkulær resonnering blir antakelsene og konklusjonene så travelt opptatte med å forfølge hverandres haler at ingen av dem bevarer en rimelig relasjon til virkeligheten.
En vanlig respons på kritikk mot tvillingmetoden er å vise til høye heritabilitetsestimater også fra familie og adopsjonsstudier. Men dette er heller en unngåelse av kritikken enn et svar på den. Selv ledende genforskere mener at familiestudier er uegnet til å skille mellom genetikk og miljø, og at adopsjonsstudier er beheftet med en rekke svakheter, som miljøpåvirkning før adopsjonen og selektiv plassering i fosterhjem. Den sterkeste adoptivmetoden er studien av eneggede tvillinger som har vokst opp hver for seg, men ingen slike studier er gjennomført for psykoser. Fordi de klassiske tvillingstudiene utgjør atferdsgenetikkens uovertrufne, sentrale informasjonskilde, følger det at hvis metoden ikke er holdbar, er hypotesen om sterke genetiske effekter ved psykoser kritisk svekket.
De høye heritabilitetsestimatene fra tvillingstudiene utgjør det sterkeste evidensgrunnlaget for påstanden om betydelige geneffekter ved psykoser. Den uholdbare grunnantakelsen om at miljøet ikke har noen betydning i tvillingdesignen, og derfor kan ses bort fra, tilsier at metoden ikke har gitt oss meningsfull kunnskap om genetiske bidrag til psykoser. Når EEA og dermed også tvillingmetoden faller, er det lite empirisk støtte igjen til teorien om at psykoser har biologiske årsaker. I stedet peker forskningen mot en dominerende betydning av sosiale og relasjonelle forhold (Read et al., 2009). Det er på tide at dette nå tas på alvor av alle. Det bør bli slutt på at man betrakter mennesker med psykoser som bærere av biologiske defekter. Dette er både stigmatiserende, devaluerende og uholdbart om mennesker som lider. I stedet bør vi bry oss om hva disse menneskene har opplevd i livet. Vi bør bruke kunnskapen om følgene av traumatisering til å skreddersy behandlingsopplegg og øke innsatsen for å forebygge at barn vokser opp i belastende miljøer.
ILLUSTRASJON AV HERITABILITSESTIMERING I TVILLINGSTUDIER |
|---|
Anta at en tvillingstudie har funnet en korrelasjon for psykoser på r = 0,50 for eneggede tvillinger og 0,15 for toeggede. Heritabilitet vil da si hvor mye genetikk betyr for den økte korrelasjonen på psykoser hos eneggede sammenliknet med toeggede. I tvillingmetoden antas likheten på et trekk for tvillingparene å skyldes deres genetiske likhet (G) og miljømessige likhet (M) (tvillingforskere ser som regel bort fra samspillet mellom genetikk og miljø). Som et utgangspunkt kan korrelasjonen på psykoser for eneggede tvillinger betraktes som summen av bidragene fra delt genetikk og delt miljø, og skrives som 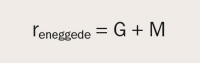
For å komme frem til et uttrykk for likheten på psykose for toeggede trekkes det veksler på at toeggede tvillinger er halvparten så like genetisk som eneggede, og på antakelsen om at eneggede par og toeggede par erfarer like korrelerte miljøer (EEA). «M» kan dermed betraktes som det samme for de to tvillingtypene, slik at trekklikheten for toeggede kan skrives som 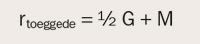
Et uttrykk for heritabilitet kan nå finnes ved å trekke korrelasjonen for toeggede fra korrelasjonen for eneggede, løse likningene for G og likestille G med heritabilitet (h) (se Suhay & Kalmoe, 2010). Resultatet blir: 
Dette betegnes som Falconers formel, som er mye brukt i estimeringen av heritabilitet (Falconer og MacKay, 1996). Hele forskjellen i de to tvillingparenes korrelasjon på trekket tilskrives her genetikk; genbidraget settes sågar lik det dobbelte av denne forskjellen. Legg merke til at miljøbidraget er forsvunnet, noe som skyldes bruken av EEA. Med en korrelasjon for psykoser på 0,50 for eneggede og 0,15 for toeggede tvillinger estimerer Falconers formel heritabilitet til 0,7 eller 70 %. |
KRITIKKEN AV TVILLINGSTUDIER I ET NØTTESKALL |
|---|

ILLUSTRASJON: KRISTIN BERG JOHNSEN
|
Teksten sto på trykk første gang i Tidsskrift for Norsk psykologforening, Vol 50, nummer 11, 2013, side 1089-1096
Referanser
Alemany, S., Goldberg, X., van Winkel, R., Gasto, C., Peralta, V. & Fañanás, L. (2012). Childhood adversity and psychosis: Examining whether the association is due to genetic confounding using a monozygotic twin differences approach. European Psychiatry, [Epub ahead of print]. http://dx.doi. org/10.1016/j.eurpsy.2012.03.001
Arseneault, L., Cannon, M., Fisher, H. L., Polanczyk, G., Moffitt, T. E. & Caspi, A. (2011). Childhood trauma and children’s emerging psychotic symptoms: a genetically sensitive longitudinal cohort study. American journal of Psychiatry, 168, 1, 65–72.
Ball, H. A., Arseneault, L., Taylor, A., Maughan, B., Caspi, C. & Moffitt, T. E. (2008). Genetic and environmental influences on victims, bullies and bully-victims in childhood. Journal of Child Psychology and Psychiatry, 49, 104–112.
Boks, M. P. (2012). Progress in gene environment studies. Biological Psychiatry, 72, 799–800.
Bouchard, T. J., Jr., Lykken, D. T., McGue, M., Segal, N. L. & Tellegen, A. (1990). Sources of human psychological differences: The Minnesota Study of Twins Reared Apart. Science, 250, 223–228.
Bouchard, T. J. & McGue, M. (2003). Genetic and environmental influences on human psychological differences. Journal of Neurobiology, 54, 4–45.
Cesarini, D., Johannesson, M., Wallace, B. & Lichtenstein, P. (2009). Heritability of overconfidence. Journal of the European Economic Association, 7, 617–627.
Champagne, F. A. & Curley, J. P. (2009). Epigenetic mechanisms mediating the long-term effects of maternal care on development. Neuroscience and Biobehavioral Reviews, 33, 593–600.
Dinwiddie, S., Heath, A. C., Dunne, M. P., Bucholz, K. K., Madden, P. A., Slutske, W. S. et al. (2000). Early sexual abuse and lifetime psychopathology: a co-twin-control study. Psychological Medicine, 30, 41–52.
Falconer, D. S. & MacKay, T. F. C. (1996). Introduction to quantitative genetics. Essex: Longmans Green.
Gershon, E. S., Alliey-Rodriguez, N. & Liu, C. (2011). After GWAS: searching for genetic risk for schizophrenia and bipolar disorder. American Journal of Psychiatry, 168, 253–256.
Horwitz, A. V., Videon, T. M., Schmitz, M. F. & Davis, D. (2003). Rethinking twins and environments: possible social sources for assumed genetic influences in twin research. Journal of Health and Social Behavior, 44, 111–129.
Jaffee, S. R., Caspi, A., Moffitt, T. E., Polo-Thomas, M., Price, T. S. & Taylor, A. (2004). The limits of child effects: Evidence for genetically mediated child effects on corporal punishment but not on physical maltreatment. Developmental Psychology, 40, 1047–1058.
Jaffee, S. R. & Price, T. S. (2008). Genotype-environment correlations: implications for determining the relationship between environmental exposures and psychiatric illness. Psychiatry, 7, 496–499.
Joseph, J. (2012). The «missing heritability» of psychiatric disorders: elusive genes or non-existent genes. Applied Developmental Sciences, 16, 65–83.
Joseph, J. (2006). The missing gene: Psychiatry, heredity and the fruitless search for genes. New York: Algora.
Kendler, K. S., MacLean, C., Neale, M., Kessler, R., Heath, A. & Eaves, L. (1991). The genetic epidemiology of bulimia nervosa. American Journal of Psychiatry, 148, 1627–1637.
Kim, Y., Zerwas, S., Trace, S. E. & Sullivan, P. F. (2011). Schizophrenia genetics: where next? Schizophrenia Bulletin, 37, 456–463.
Kringlen, E. (1967). Heredity and environment in the functional psychoses: an epidemiological clinical study. Oslo: Universitetsforlaget.
Price, T. S. & Jaffee, S. R. (2008). Effects of the family environment: Gene–environment interaction and passive gene–environment correlation. Developmental Psychology, 44, 305–315.
Read, J., Bentall, R. P. & Fosse, R. (2009). Time to abandon the bio-bio-bio model of psychosis: exploring the epigenetic and psychosocial mechanisms by which adverse life events lead to psychotic symptoms. Epidemiologia e Psichiatria Sociale, 18, 299–310.
Ross, C. A. (2013). The equal environments assumption in schizophrenia genetics. Psychosis, in press.
Scarr, S. & Carter-Saltzman, L. (1979). Twin method: Defense of a critical assumption. Behavior Genetics, 9, 527–542.
Shevlin, M., Dorahy, M. J. & Adamson, G. (2007). Trauma and psychosis: an analysis of the National Comorbidity Survey. American journal of Psychiatry, 164, 166–169.
Stein, M. B., Jang, K. L., Taylor, S., Vernon, P. A. & Livesley, W. J. (2002). Genetic and environmental influences on trauma exposure and posttraumatic stress disorder symptoms: a twin study. American journal of Psychiatry, 159, 1675–1681.
Suhay, E. & Kalmoe, N. P. (2010). Violations of the equal environment assumption in twin studies of political traits. http://sites.lafayette.edu/suhaye/files/2010/01/Violations-of-the-EEA-Su...
The International Schizophrenia Consortium (2008). Rare chromosomal deletions and duplications increase risk of schizophrenia. Nature, 455, 237–241.
Varese, F., Smeets, F., Drukker, M., Lieverse, R., Lataster, T., Viechtbauer, W. et al. (2012). Childhood adversities increase the risk of psychosis: A meta-analysis of patient-control, prospective- and cross-sectional cohort studies. Schizophrenia Bulletin, 38, 661–671.


Kommenter denne artikkelen